
 |
 |
 |
 |
 |
 |
 |
 |
Kookaburras
genus DaceloKookaburras (genus Dacelo) are large to very large (total length 28-42 cm/11-17 in) terrestrial kingfishers native to Australia and New Guinea, the name a loanword from Wiradjuri guuguubarra, which is onomatopoeic of its call. The single member of the genus Clytoceyx, while commonly referred to as the Shovel-billed Kookaburra, is not treated in this article.
Kookaburras are best known for their unmistakable call, which is uncannily like loud, echoing human laughter — good-natured, but rather hysterical, merriment in the case of the well-known Laughing Kookaburra (Dacelo novaeguineae); and maniacal cackling in the case of the slightly smaller Blue-winged Kookaburra (D. leachii). They are generally not closely associated with water, and can be found in habitats ranging from humid forest to arid savanna, but also in suburban and residential areas near running water and where food can be searched for easily.
Classification and species
There are four known species of kookaburras found in Australia, New Guinea, and the Aru Islands.
Unusual for close relatives, the Laughing and Blue-winged species are direct competitors in the area where their ranges overlap. This suggests that the two species, though having common stock, evolved in isolation (possibly during a period when Australia and New Guinea were more distant — see Australia-New Guinea) and were only brought back into contact in relatively recent geological times.
Species
All kookaburras are sexually dimorphic, but this is only obvious in the Blue-winged and the Rufous-bellied, where males have blue tails, females rufous.
* Rufous-bellied Kookaburra (Dacelo gaudichaud).
* Spangled Kookaburra (Dacelo tyro).
* Blue-winged Kookaburra (Dacelo leachii).
* Laughing Kookaburra (Dacelo novaeguineae).Behaviour
Kookaburras are carnivorous. They will eat lizards, snakes, insects, mice, other small birds, and raw meat. The most social birds will accept handouts from humans and will take raw or cooked meat (even if at high temperature) from on or near open-air barbecues left unattended. It is generally not advised to feed the birds too regularly as meat alone does not include calcium and other nutrients essential to the bird. Remainders of mince on the bird's beak can fester and cause problems for the bird.
They are territorial, and often live with the partly grown chicks of the previous season. They often sing as a chorus to mark their territory.
In the wild, kookaburras are known to eat babies of other birds and snakes, and insects and small reptiles and even other small birds, such as finches if they are lucky enough to catch them. In zoos, they are usually fed food for birds of prey, and dead baby chicks.
Web Reference: http://en.wikipedia.org/wiki/Kookaburra
Birds
Fossil range: 150-0 Ma
Late Jurassic - Recent
Scientific classification
Superdomain: Neomura
Domain: Eukaryota
Kingdom: Animalia
Subkingdom: Eumetazoa
Superphylum: Deuterostomia
Phylum: Chordata
Subphylum: Vertebrata
Infraphylum: Tetrapoda
Microphylum: Amniota
Nanophylum: Diapsida
(unranked): Archosauria
Superclass: Archosauromorpha
Class: Aves (Linnaeus, 1758)Birds (class Aves) are bipedal, endothermic (warm-blooded), vertebrate animals that lay eggs. There are around 10,000 living species, making them the most numerous tetrapod vertebrates. They inhabit ecosystems across the globe, from the Arctic to the Antarctic. Birds range in size from the 5 cm (2 in) Bee Hummingbird to the 2.7 m (9 ft) Ostrich. The fossil record indicates that birds evolved from theropod dinosaurs during the Jurassic period, around 150-200 Ma (million years ago), and the earliest known bird is the Late Jurassic Archaeopteryx, c 155-150 Ma. Most paleontologists regard birds as the only clade of dinosaurs that survived the Cretaceous-Tertiary extinction event approximately 65.5 Ma.
Modern birds are characterised by feathers, a beak with no teeth, the laying of hard-shelled eggs, a high metabolic rate, a four-chambered heart, and a lightweight but strong skeleton. All birds have forelimbs modified as wings and most can fly, with some exceptions including ratites, penguins, and a number of diverse endemic island species. Birds also have unique digestive and respiratory systems that are highly adapted for flight. Some birds, especially corvids and parrots, are among the most intelligent animal species; a number of bird species have been observed manufacturing and using tools, and many social species exhibit cultural transmission of knowledge across generations.
Many species undertake long distance annual migrations, and many more perform shorter irregular movements. Birds are social; they communicate using visual signals and through calls and songs, and participate in social behaviours including cooperative breeding and hunting, flocking, and mobbing of predators. The vast majority of bird species are socially monogamous, usually for one breeding season at a time, sometimes for years, but rarely for life. Other species have breeding systems that are polygynous ("many females") or, rarely, polyandrous ("many males"). Eggs are usually laid in a nest and incubated by the parents. Most birds have an extended period of parental care after hatching.
Many species are of economic importance, mostly as sources of food acquired through hunting or farming. Some species, particularly songbirds and parrots, are popular as pets. Other uses include the harvesting of guano (droppings) for use as a fertiliser. Birds figure prominently in all aspects of human culture from religion to poetry to popular music. About 120-130 species have become extinct as a result of human activity since the 17th century, and hundreds more before then. Currently about 1,200 species of birds are threatened with extinction by human activities, though efforts are underway to protect them.
Evolution and taxonomy
The first classification of birds was developed by Francis Willughby and John Ray in their 1676 volume Ornithologiae. Carolus Linnaeus modified that work in 1758 to devise the taxonomic classification system currently in use. Birds are categorised as the biological class Aves in Linnaean taxonomy. Phylogenetic taxonomy places Aves in the dinosaur clade Theropoda. Aves and a sister group, the clade Crocodilia, together are the sole living members of the reptile clade Archosauria. Phylogenetically, Aves is commonly defined as all descendants of the most recent common ancestor of modern birds and Archaeopteryx lithographica. Archaeopteryx, from the Tithonian stage of the Late Jurassic (some 150-145 million years ago), is the earliest known bird under this definition. Others, including Jacques Gauthier and adherents of the Phylocode system, have defined Aves to include only the modern bird groups, excluding most groups known only from fossils, and assigning them, instead, to the Avialae in part to avoid the uncertainties about the placement of Archaeopteryx in relation to animals traditionally thought of as theropod dinosaurs.
All modern birds lie within the subclass Neornithes, which has two subdivisions: the Paleognathae, containing mostly flightless birds like ostriches, and the wildly diverse Neognathae, containing all other birds. These two subdivisions are often given the rank of superorder, although Livezey & Zusi assigned them "cohort" rank. Depending on the taxonomic viewpoint, the number of known living bird species varies anywhere from 9,800 to 10,050.
Dinosaurs and the origin of birds
Fossil evidence and intensive biological analyses have demonstrated beyond any reasonable doubt that birds are theropod dinosaurs. More specifically, they are members of Maniraptora, a group of theropods which includes dromaeosaurs and oviraptorids, among others. As scientists discover more non-avian theropods that are closely related to birds, the previously clear distinction between non-birds and birds has become blurred. Recent discoveries in the Liaoning Province of northeast China, which demonstrate that many small theropod dinosaurs had feathers, contribute to this ambiguity.
The consensus view in contemporary paleontology is that the birds, Aves, are the closest relatives of the deinonychosaurs, which include dromaeosaurids and troodontids. Together, these three form a group called Paraves. The basal dromaeosaur Microraptor has features which may have enabled it to glide or fly. The most basal deinonychosaurs are very small. This evidence raises the possibility that the ancestor of all paravians may have been arboreal, and/or may have been able to glide.
The Late Jurassic Archaeopteryx is well-known as one of the first transitional fossils to be found and it provided support for the theory of evolution in the late 19th century. Archaeopteryx has clearly reptilian characters: teeth, clawed fingers, and a long, lizard-like tail, but it has finely preserved wings with flight feathers identical to those of modern birds. It is not considered a direct ancestor of modern birds, but is the oldest and most primitive member of Aves or Avialae, and it is probably closely related to the real ancestor. It has even been suggested that Archaeopteryx was a dinosaur that was no more closely related to birds than were other dinosaur groups, and that Avimimus was more likely to be the ancestor of all birds than Archaeopteryx.
Alternative theories and controversies
There have been many controversies in the study of the origin of birds. Early disagreements included whether birds evolved from dinosaurs or more primitive archosaurs. Within the dinosaur camp there were disagreements as to whether ornithischian or theropod dinosaurs were the more likely ancestors. Although ornithischian (bird-hipped) dinosaurs share the hip structure of modern birds, birds are thought to have originated from the saurischian (lizard-hipped) dinosaurs, and therefore evolved their hip structure independently. In fact, a bird-like hip structure evolved a third time among a peculiar group of theropods known as the Therizinosauridae. A few scientists suggest that birds are not dinosaurs, but evolved from early archosaurs like Longisquama.
Early evolution of birds
Aves
Archaeopteryx
Pygostylia
Confuciusornithidae
Ornithothoraces
Enantiornithes
Ornithurae
Hesperornithiformes
NeornithesBirds diversified into a wide variety of forms during the Cretaceous Period. Many groups retained primitive characteristics, such as clawed wings and teeth, though the latter were lost independently in a number of bird groups, including modern birds (Neornithes). While the earliest forms, such as Archaeopteryx and Jeholornis, retained the long bony tails of their ancestors, the tails of more advanced birds were shortened with the advent of the pygostyle bone in the clade Pygostylia.
The first large, diverse lineage of short-tailed birds to evolve were the Enantiornithes, or "opposite birds", so named because the construction of their shoulder bones was in reverse to that of modern birds. Enantiornithes occupied a wide array of ecological niches, from sand-probing shorebirds and fish-eaters to tree-dwelling forms and seed-eaters. More advanced lineages also specialised in eating fish, like the superficially gull-like subclass of Ichthyornithes ("fish birds"). One order of Mesozoic seabirds, the Hesperornithiformes, became so well adapted to hunting fish in marine environments that they lost the ability to fly and became primarily aquatic. Despite their extreme specialisations, the Hesperornithiformes represent some of the closest relatives of modern birds.
Radiation of modern birds
Containing all modern birds, the subclass Neornithes is, due to the discovery of Vegavis, now known to have evolved into some basic lineages by the end of the Cretaceous and is split into two superorders, the Paleognathae and Neognathae. The paleognaths include the tinamous of Central and South America and the ratites. The basal divergence from the remaining Neognathes was that of the Galloanserae, the superorder containing the Anseriformes (ducks, geese, swans and screamers) and the Galliformes (the pheasants, grouse, and their allies, together with the mound builders and the guans and their allies). The dates for the splits are much debated by scientists. It is agreed that the Neornithes evolved in the Cretaceous, and that the split between the Galloanseri from other Neognathes occurred before the K-T extinction event, but there are different opinions about whether the radiation of the remaining Neognathes occurred before or after the extinction of the other dinosaurs. This disagreement is in part caused by a divergence in the evidence; molecular dating suggests a Cretaceous radiation, while fossil evidence supports a Tertiary radiation. Attempts to reconcile the molecular and fossil evidence have proved controversial.
The classification of birds is a contentious issue. Sibley and Ahlquist's Phylogeny and Classification of Birds (1990) is a landmark work on the classification of birds, although it is frequently debated and constantly revised. Most evidence seems to suggest that the assignment of orders is accurate, but scientists disagree about the relationships between the orders themselves; evidence from modern bird anatomy, fossils and DNA have all been brought to bear on the problem, but no strong consensus has emerged. More recently, new fossil and molecular evidence is providing an increasingly clear picture of the evolution of modern bird orders.
Modern bird orders: Classification
Neornithes
Paleognathae
Struthioniformes
Tinamiformes
Neognathae
Other birds
Galloanserae
Anseriformes
GalliformesBasal divergences of modern birds based on Sibley-Ahlquist taxonomy
This is a list of the taxonomic orders in the subclass Neornithes, or modern birds. This list uses the traditional classification (the so-called Clements order), revised by the Sibley-Monroe classification. The list of birds gives a more detailed summary of the orders, including families.
Subclass Neornithes
Paleognathae:* Struthioniformes-ostriches, emus, kiwis, and allies
* Tinamiformes-tinamousNeognathae:
* Anseriformes-waterfowl
* Galliformes-fowl
* Charadriiformes-gulls, button-quails, plovers and allies
* Gaviiformes-loons
* Podicipediformes-grebes
* Procellariiformes-albatrosses, petrels, and allies
* Sphenisciformes-penguins
* Pelecaniformes-pelicans and allies
* Phaethontiformes-tropicbirds
* Ciconiiformes-storks and allies
* Cathartiformes-New World vultures
* Phoenicopteriformes-flamingos
* Falconiformes-falcons, eagles, hawks and allies
* Gruiformes-cranes and allies
* Pteroclidiformes-sandgrouse
* Columbiformes-doves and pigeons
* Psittaciformes-parrots and allies
* Cuculiformes-cuckoos and turacos
* Opisthocomiformes-hoatzin
* Strigiformes-owls
* Caprimulgiformes-nightjars and allies
* Apodiformes-swifts and hummingbirds
* Coraciiformes-kingfishers and allies
* Piciformes-woodpeckers and allies
* Trogoniformes-trogons
* Coliiformes-mousebirds
* Passeriformes-passerinesThe radically different Sibley-Monroe classification (Sibley-Ahlquist taxonomy), based on molecular data, found widespread adoption in a few aspects, as recent molecular, fossil, and anatomical evidence supported the Galloanserae for example.
Distribution
Birds live and breed in most terrestrial habitats and on all seven continents, reaching their southern extreme in the Snow Petrel's breeding colonies up to 440 kilometres (270 mi) inland in Antarctica. The highest bird diversity occurs in tropical regions. It was earlier thought that this high diversity was the result of higher speciation rates in the tropics, however recent studies found higher speciation rates in the high latitudes that were offset by greater extinction rates than in the tropics. Several families of birds have adapted to life both on the world's oceans and in them, with some seabird species coming ashore only to breed and some penguins have been recorded diving up to 300 metres (980 ft).
Many bird species have established breeding populations in areas to which they have been introduced by humans. Some of these introductions have been deliberate; the Ring-necked Pheasant, for example, has been introduced around the world as a game bird. Others have been accidental, such as the establishment of wild Monk Parakeets in several North American cities after their escape from captivity. Some species, including Cattle Egret, Yellow-headed Caracara and Galah, have spread naturally far beyond their original ranges as agricultural practices created suitable new habitat.
Anatomy and physiology
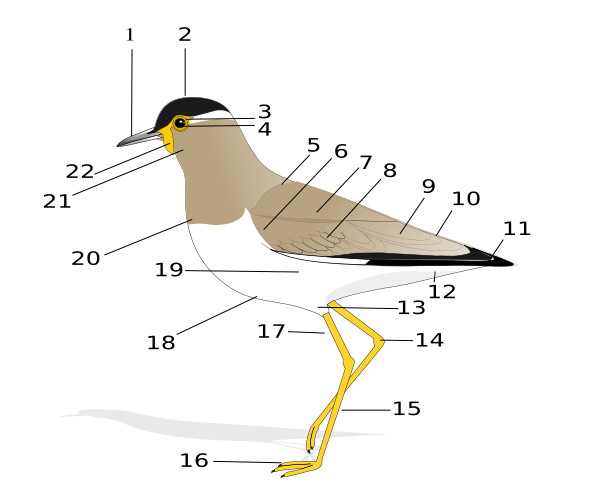
External anatomy of a bird: 1 Beak, 2 Head, 3 Iris, 4 Pupil, 5 Mantle, 6 Lesser coverts, 7 Scapulars, 8 Median coverts, 9 Tertials, 10 Rump, 11 Primaries, 12 Vent, 13 Thigh, 14 Tibio-tarsal articulation, 15 Tarsus, 16 Feet, 17 Tibia, 18 Belly, 19 Flanks, 20 Breast, 21 Throat, 22 Wattle
Compared with other vertebrates, birds have a body plan that shows many unusual adaptations, mostly to facilitate flight.
The skeleton consists of very lightweight bones. They have large air-filled cavities (called pneumatic cavities) which connect with the respiratory system. The skull bones are fused and do not show cranial sutures. The orbits are large and separated by a bony septum. The spine has cervical, thoracic, lumbar and caudal regions with the number of cervical (neck) vertebrae highly variable and especially flexible, but movement is reduced in the anterior thoracic vertebrae and absent in the later vertebrae. The last few are fused with the pelvis to form the synsacrum. The ribs are flattened and the sternum is keeled for the attachment of flight muscles except in the flightless bird orders. The forelimbs are modified into wings.
Like the reptiles, birds are primarily uricotelic, that is, their kidneys extract nitrogenous wastes from their bloodstream and excrete it as uric acid instead of urea or ammonia via the ureters into the intestine. Birds do not have a urinary bladder or external urethral opening and uric acid is excreted along with feces as a semisolid waste. However, birds such as hummingbirds can be facultatively ammonotelic, excreting most of the nitrogenous wastes as ammonia. They also excrete creatine, rather than creatinine like mammals. This material, as well as the output of the intestines, emerges from the bird's cloaca. The cloaca is a multi-purpose opening: waste is expelled through it, birds mate by joining cloaca, and females lay eggs from it. In addition, many species of birds regurgitate pellets. The digestive system of birds is unique, with a crop for storage and a gizzard that contains swallowed stones for grinding food to compensate for the lack of teeth. Most birds are highly adapted for rapid digestion to aid with flight. Some migratory birds have adapted to use protein from many parts of their bodies, including protein from the intestines, as additional energy during migration.
Birds have one of the most complex respiratory systems of all animal groups. Upon inhalation, 75% of the fresh air bypasses the lungs and flows directly into a posterior air sac which extends from the lungs and connects with air spaces in the bones and fills them with air. The other 25% of the air goes directly into the lungs. When the bird exhales, the used air flows out of the lung and the stored fresh air from the posterior air sac is simultaneously forced into the lungs. Thus, a bird's lungs receive a constant supply of fresh air during both inhalation and exhalation. Sound production is achieved using the syrinx, a muscular chamber with several tympanic membranes which is situated at the lower end of the trachea, from where it separates. The bird's heart has four chambers and the right aortic arch gives rise to systemic circulation (unlike in the mammals where the left arch is involved). The postcava receives blood from the limbs via the renal portal system. Unlike in mammals, the red blood cells in birds have a nucleus.
The nervous system is large relative to the bird's size. The most developed part of the brain is the one that controls the flight-related functions, while the cerebellum coordinates movement and the cerebrum controls behaviour patterns, navigation, mating and nest building. Most birds have a poor sense of smell with notable exceptions including kiwis, New World vultures and tubenoses. The avian visual system is usually highly developed. Water birds have special flexible lenses, allowing accommodation for vision in air and water. Some species also have dual fovea. Birds are tetrachromatic, possessing ultraviolet (UV) sensitive cone cells in the eye as well as green, red and blue ones. This allows them to perceive ultraviolet light, which is involved in courtship. Many birds show plumage patterns in ultraviolet that are invisible to the human eye; some birds whose sexes appear similar to the naked eye are distinguished by the presence of ultraviolet reflective patches on their feathers. Male Blue Tits have an ultraviolet reflective crown patch which is displayed in courtship by posturing and raising of their nape feathers. Ultraviolet light is also used in foraging-kestrels have been shown to search for prey by detecting the UV reflective urine trail marks left on the ground by rodents. The eyelids of a bird are not used in blinking. Instead the eye is lubricated by the nictitating membrane, a third eyelid that moves horizontally. The nictitating membrane also covers the eye and acts as a contact lens in many aquatic birds. The bird retina has a fan shaped blood supply system called the pecten. Most birds cannot move their eyes, although there are exceptions, such as the Great Cormorant. Birds with eyes on the sides of their heads have a wide visual field, while birds with eyes on the front of their heads, such as owls, have binocular vision and can estimate the depth of field. The avian ear lacks external pinnae but is covered by feathers, although in some birds, such as the Asio, Bubo and Otus owls, these feathers form tufts which resemble ears. The inner ear has a cochlea, but it is not spiral as in mammals.
A few species are able to use chemical defenses against predators; some Procellariiformes can eject an unpleasant oil against an aggressor, and some species of pitohuis from New Guinea have a powerful neurotoxin in their skin and feathers.
Chromosomes
Birds have two sexes: male and female. The sex of birds is determined by the Z and W sex chromosomes, rather than by the X and Y chromosomes present in mammals. Male birds have two Z chromosomes (ZZ), and female birds have a W chromosome and a Z chromosome (WZ).
In nearly all species of birds, an individual's sex is determined at fertilization. However, one recent study demonstrated temperature-dependent sex determination among Australian Brush-turkeys, for which higher temperatures during incubation resulted in a higher female-to-male sex ratio.
Feathers, plumage, and scales
Feathers are a feature unique to birds. They facilitate flight, provide insulation that aids in thermoregulation, and are used in display, camouflage, and signaling. There are several types of feathers, each serving its own set of purposes. Feathers are epidermal growths attached to the skin and arise only in specific tracts of skin called pterylae. The distribution pattern of these feather tracts (pterylosis) is used in taxonomy and systematics. The arrangement and appearance of feathers on the body, called plumage, may vary within species by age, social status, and sex.
Plumage is regularly moulted; the standard plumage of a bird that has moulted after breeding is known as the "non-breeding" plumage, or - in the Humphrey-Parkes terminology - "basic" plumage; breeding plumages or variations of the basic plumage are known under the Humphrey-Parkes system as "alternate" plumages. Moulting is annual in most species, although some may have two moults a year, and large birds of prey may moult only once every few years. Moulting patterns vary across species. In passerines, flight feathers are replaced one at a time with the innermost primary being the first. When the fifth of sixth primary is replaced, the outermost tertiaries begin to drop. After the innermost tertiaries are moulted, the secondaries starting from the innermost begin to drop and this proceeds to the outer feathers (centrifugal moult). The greater primary coverts are moulted in synchrony with the primary that they overlap. A small number of species, such as ducks and geese, lose all of their flight feathers at once, temporarily becoming flightless. As a general rule, the tail feathers are moulted and replaced starting with the innermost pair. Centripetal moults of tail feathers are however seen in the Phasianidae. The centrifugal moult is modified in the tail feathers of woodpeckers and treecreepers, in that it begins with the second innermost pair of feathers and finishes with the central pair of feathers so that the bird maintains a functional climbing tail. The general pattern seen in passerines is that the primaries are replaced outward, secondaries inward, and the tail from center outward. Before nesting, the females of most bird species gain a bare brood patch by losing feathers close to the belly. The skin there is well supplied with blood vessels and helps the bird in incubation.
Feathers require maintenance and birds preen or groom them daily, spending an average of around 9% of their daily time on this. The bill is used to brush away foreign particles and to apply waxy secretions from the uropygial gland; these secretions protect the feathers' flexibility and act as an antimicrobial agent, inhibiting the growth of feather-degrading bacteria. This may be supplemented with the secretions of formic acid from ants, which birds receive through a behaviour known as anting, to remove feather parasites.
The scales of birds are composed of the same keratin as beaks, claws, and spurs. They are found mainly on the toes and metatarsus, but may be found further up on the ankle in some birds. Most bird scales do not overlap significantly, except in the cases of kingfishers and woodpeckers. The scales of birds are thought to be homologous to those of reptiles and mammals.
Flight
Most birds can fly, which distinguishes them from almost all other vertebrates. Flight is the primary means of locomotion for most bird species and is used for breeding, feeding, and predator avoidance and escape. Birds have various adaptations for flight, including a lightweight skeleton, two large flight muscles (the pectoralis-accounting for 15% of the total mass of the bird-and the supracoracoideus), and a modified forelimb (wing) that serves as an aerofoil. Wing shape and size generally determine a bird species' type of flight; many birds combine powered, flapping flight with less energy-intensive soaring flight. About 60 extant bird species are flightless, as were many extinct birds. Flightlessness often arises in birds on isolated islands, probably due to limited resources and the absence of land predators. Though flightless, penguins use similar musculature and movements to "fly" through the water, as do auks, shearwaters and dippers.
Behaviour
Most birds are diurnal, but some birds, such as many species of owls and nightjars, are nocturnal or crepuscular (active during twilight hours), and many coastal waders feed when the tides are appropriate, by day or night.
Diet and feeding
Birds' diets are varied and often include nectar, fruit, plants, seeds, carrion, and various small animals, including other birds. Because birds have no teeth, their digestive system is adapted to process unmasticated food items that are swallowed whole.
Birds that employ many strategies to obtain food or feed on a variety of food items are called generalists, while others that concentrate time and effort on specific food items or have a single strategy to obtain food are considered specialists. Birds' feeding strategies vary by species. Many birds glean for insects, invertebrates, fruit, or seeds. Some hunt insects by suddenly attacking from a branch. Nectar feeders such as hummingbirds, sunbirds, lories, and lorikeets amongst others have specially adapted brushy tongues and in many cases bills designed to fit co-adapted flowers. Kiwis and shorebirds with long bills probe for invertebrates; shorebirds' varied bill lengths and feeding methods result in the separation of ecological niches. Loons, diving ducks, penguins and auks pursue their prey underwater, using their wings or feet for propulsion, while aerial predators such as sulids, kingfishers and terns plunge dive after their prey. Flamingos, three species of prion, and some ducks are filter feeders. Geese and dabbling ducks are primarily grazers. Some species, including frigatebirds, gulls, and skuas, engage in kleptoparasitism, stealing food items from other birds. Kleptoparasitism is thought to be a supplement to food obtained by hunting, rather than a significant part of any species' diet; a study of Great Frigatebirds stealing from Masked Boobies estimated that the frigatebirds stole at most 40% of their food and on average stole only 5%. Other birds are scavengers; some of these, like vultures, are specialised carrion eaters, while others, like gulls, corvids, or other birds of prey, are opportunists.
Water and drinking
Water is needed by many birds although their mode of excretion and lack of sweat glands reduces the physiological demands. Some desert birds can obtain their water needs entirely from moisture in their food. They may also have other adaptations such as allowing their body temperature to rise, saving on moisture loss from evaporative cooling or panting. Seabirds can drink seawater and have salt glands inside the head that eliminate excess salt out of the nostrils.
Most birds scoop water in their beaks and raise their head to let water run down the throat. Some species, especially of arid zones, belonging to the pigeon, finch, mousebird, button-quail and bustard families are capable of sucking up water without the need to tilt back their heads. Some desert birds depend on water sources and sandgrouse are particularly well-known for their daily congregations at waterholes. Nesting sandgrouse carry water to their young by wetting their belly feathers.
Migration
Many bird species migrate to take advantage of global differences of seasonal temperatures, therefore optimising availability of food sources and breeding habitat. These migrations vary among the different groups. Many landbirds, shorebirds, and waterbirds undertake annual long distance migrations, usually triggered by the length of daylight as well as weather conditions. These birds are characterised by a breeding season spent in the temperate or arctic/antarctic regions and a non-breeding season in the tropical regions or opposite hemisphere. Before migration, birds substantially increase body fats and reserves and reduce the size of some of their organs. Migration is highly demanding energetically, particularly as birds need to cross deserts and oceans without refuelling. Landbirds have a flight range of around 2,500 km (1,600 mi) and shorebirds can fly up to 4,000 km (2,500 mi), although the Bar-tailed Godwit is capable of non-stop flights of up to 10,200 km (6,300 mi). Seabirds also undertake long migrations, the longest annual migration being those of Sooty Shearwaters, which nest in New Zealand and Chile and spend the northern summer feeding in the North Pacific off Japan, Alaska and California, an annual round trip of 64,000 km (39,800 mi). Other seabirds disperse after breeding, travelling widely but having no set migration route. Albatrosses nesting in the Southern Ocean often undertake circumpolar trips between breeding seasons.
Some bird species undertake shorter migrations, travelling only as far as is required to avoid bad weather or obtain food. Irruptive species such as the boreal finches are one such group and can commonly be found at a location in one year and absent the next. This type of migration is normally associated with food availability. Species may also travel shorter distances over part of their range, with individuals from higher latitudes travelling into the existing range of conspecifics; others undertake partial migrations, where only a fraction of the population, usually females and subdominant males, migrates. Partial migration can form a large percentage of the migration behaviour of birds in some regions; in Australia, surveys found that 44% of non-passerine birds and 32% of passerines were partially migratory. Altitudinal migration is a form of short distance migration in which birds spend the breeding season at higher altitudes elevations and move to lower ones during suboptimal conditions. It is most often triggered by temperature changes and usually occurs when the normal territories also become inhospitable due to lack of food. Some species may also be nomadic, holding no fixed territory and moving according to weather and food availability. Parrots as a family are overwhelmingly neither migratory nor sedentary but considered to either be dispersive, irruptive, nomadic or undertake small and irregular migrations.
The ability of birds to return to precise locations across vast distances has been known for some time; in an experiment conducted in the 1950s a Manx Shearwater released in Boston returned to its colony in Skomer, Wales within 13 days, a distance of 5,150 km (3,200 mi). Birds navigate during migration using a variety of methods. For diurnal migrants, the sun is used to navigate by day, and a stellar compass is used at night. Birds that use the sun compensate for the changing position of the sun during the day by the use of an internal clock. Orientation with the stellar compass depends on the position of the constellations surrounding Polaris. These are backed up in some species by their ability to sense the Earth's geomagnetism through specialised photoreceptors.
Communication
The startling display of the Sunbittern mimics a large predator.Birds communicate using primarily visual and auditory signals. Signals can be interspecific (between species) and intraspecific (within species).
Birds sometimes use plumage to assess and assert social dominance, to display breeding condition in sexually selected species, or to make threatening displays, as in the Sunbittern's mimicry of a large predator to ward off hawks and protect young chicks. Variation in plumage also allows for the identification of birds, particularly between species. Visual communication among birds may also involve ritualised displays, which have developed from non-signalling actions such as preening, the adjustments of feather position, pecking, or other behaviour. These displays may signal aggression or submission or may contribute to the formation of pair-bonds. The most elaborate displays occur during courtship, where "dances" are often formed from complex combinations of many possible component movements; males' breeding success may depend on the quality of such displays.
Bird calls and songs, which are produced in the syrinx, are the major means by which birds communicate with sound. This communication can be very complex; some species can operate the two sides of the syrinx independently, allowing the simultaneous production of two different songs. Calls are used for a variety of purposes, including mate attraction, evaluation of potential mates, bond formation, the claiming and maintenance of territories, the identification of other individuals (such as when parents look for chicks in colonies or when mates reunite at the start of breeding season), and the warning of other birds of potential predators, sometimes with specific information about the nature of the threat. Some birds also use mechanical sounds for auditory communication. The Coenocorypha snipes of New Zealand drive air through their feathers, woodpeckers drum territorially, and Palm Cockatoos use tools to drum.
Flocking and other associations
While some birds are essentially territorial or live in small family groups, other birds may form large flocks. The principal benefits of flocking are safety in numbers and increased foraging efficiency. Defence against predators is particularly important in closed habitats like forests, where ambush predation is common and multiple eyes can provide a valuable early warning system. This has led to the development of many mixed-species feeding flocks, which are usually composed of small numbers of many species; these flocks provide safety in numbers but reduce potential competition for resources. Costs of flocking include bullying of socially subordinate birds by more dominant birds and the reduction of feeding efficiency in certain cases.
Birds sometimes also form associations with non-avian species. Plunge-diving seabirds associate with dolphins and tuna, which push shoaling fish towards the surface. Hornbills have a mutualistic relationship with Dwarf Mongooses, in which they forage together and warn each other of nearby birds of prey and other predators.
Resting and roosting
The high metabolic rates of birds during the active part of the day is supplemented by rest at other times. Sleeping birds often use a type of sleep known as vigilant sleep, where periods of rest are interspersed with quick eye-opening 'peeks', allowing them to be sensitive to disturbances and enable rapid escape from threats. Swifts are believed to be able to sleep in flight and radar observations suggest that they orient themselves to face the wind in their roosting flight. It has been suggested that there may be certain kinds of sleep which are possible even when in flight. Some birds have also demonstrated the capacity to fall into slow-wave sleep one hemisphere of the brain at a time. The birds tend to exercise this ability depending upon its position relative to the outside of the flock. This may allow the eye opposite the sleeping hemisphere to remain vigilant for predators by viewing the outer margins of the flock. This adaptation is also known from marine mammals. Communal roosting is common because it lowers the loss of body heat and decreases the risks associated with predators. Roosting sites are often chosen with regard to thermoregulation and safety.
Many sleeping birds bend their heads over their backs and tuck their bills in their back feathers, although others place their beaks among their breast feathers. Many birds rest on one leg, while some may pull up their legs into their feathers, especially in cold weather. Perching birds have a tendon locking mechanism that helps them hold on to the perch when they are asleep. Many ground birds, such as quails and pheasants, roost in trees. A few parrots of the genus Loriculus roost hanging upside down. Some hummingbirds go into a nightly state of torpor accompanied with a reduction of their metabolic rates. This physiological adaptation shows nearly a hundred other species, including owlet-nightjars, nightjars, and woodswallows. One species, the Common Poorwill, even enters a state of hibernation. Birds do not have sweat glands, but they may cool themselves by moving to shade, standing in water, panting, increasing their surface area, fluttering their throat or by using special behaviours like urohydrosis to cool themselves.
Breeding
Social systems
Ninety-five percent of bird species are socially monogamous. These species pair for at least the length of the breeding season or-in some cases-for several years or until the death of one mate. Monogamy allows for biparental care, which is especially important for species in which females require males' assistance for successful brood-rearing. Among many socially monogamous species, extra-pair copulation (infidelity) is common. Such behaviour typically occurs between dominant males and females paired with subordinate males, but may also be the result of forced copulation in ducks and other anatids. For females, possible benefits of extra-pair copulation include getting better genes for her offspring and insuring against the possibility of infertility in her mate. Males of species that engage in extra-pair copulations will closely guard their mates to ensure the parentage of the offspring that they raise.
Other mating systems, including polygyny, polyandry, polygamy, polygynandry, and promiscuity, also occur. Polygamous breeding systems arise when females are able to raise broods without the help of males. Some species may use more than one system depending on the circumstances.
Breeding usually involves some form of courtship display, typically performed by the male. Most displays are rather simple and involve some type of song. Some displays, however, are quite elaborate. Depending on the species, these may include wing or tail drumming, dancing, aerial flights, or communal lekking. Females are generally the ones that drive partner selection, although in the polyandrous phalaropes, this is reversed: plainer males choose brightly coloured females. Courtship feeding, billing and allopreening are commonly performed between partners, generally after the birds have paired and mated.
Territories, nesting and incubation
Many birds actively defend a territory from others of the same species during the breeding season; maintenance of territories protects the food source for their chicks. Species that are unable to defend feeding territories, such as seabirds and swifts, often breed in colonies instead; this is thought to offer protection from predators. Colonial breeders defend small nesting sites, and competition between and within species for nesting sites can be intense.
All birds lay amniotic eggs with hard shells made mostly of calcium carbonate. Hole and burrow nesting species tend to lay white or pale eggs, while open nesters lay camouflaged eggs. There are many exceptions to this pattern, however; the ground-nesting nightjars have pale eggs, and camouflage is instead provided by their plumage. Species that are victims of brood parasites have varying egg colours to improve the chances of spotting a parasite's egg, which forces female parasites to match their eggs to those of their hosts.
Bird eggs are usually laid in a nest. Most species create somewhat elaborate nests, which can be cups, domes, plates, beds scrapes, mounds, or burrows. Some bird nests, however, are extremely primitive; albatross nests are no more than a scrape on the ground. Most birds build nests in sheltered, hidden areas to avoid predation, but large or colonial birds-which are more capable of defence-may build more open nests. During nest construction, some species seek out plant matter from plants with parasite-reducing toxins to improve chick survival, and feathers are often used for nest insulation. Some bird species have no nests; the cliff-nesting Common Guillemot lays its eggs on bare rock, and male Emperor Penguins keep eggs between their body and feet. The absence of nests is especially prevalent in ground-nesting species where the newly hatched young are precocial.
Incubation, which optimises temperature for chick development, usually begins after the last egg has been laid. In monogamous species incubation duties are often shared, whereas in polygamous species one parent is wholly responsible for incubation. Warmth from parents passes to the eggs through brood patches, areas of bare skin on the abdomen or breast of the incubating birds. Incubation can be an energetically demanding process; adult albatrosses, for instance, lose as much as 83 grams (2.9 oz) of body weight per day of incubation. The warmth for the incubation of the eggs of megapodes comes from the sun, decaying vegetation or volcanic sources. Incubation periods range from 10 days (in woodpeckers, cuckoos and passerine birds) to over 80 days (in albatrosses and kiwis).
Parental care and fledging
At the time of their hatching, chicks range in development from helpless to independent, depending on their species. Helpless chicks are termed altricial, and tend to be born small, blind, immobile and naked; chicks that are mobile and feathered upon hatching are termed precocial. Altricial chicks need help thermoregulating and must be brooded for longer than precocial chicks. Chicks at neither of these extremes can be semi-precocial or semi-altricial.
The length and nature of parental care varies widely amongst different orders and species. At one extreme, parental care in megapodes ends at hatching; the newly-hatched chick digs itself out of the nest mound without parental assistance and can fend for itself immediately. At the other extreme, many seabirds have extended periods of parental care, the longest being that of the Great Frigatebird, whose chicks take up to six months to fledge and are fed by the parents for up to an additional 14 months.
In some species, both parents care for nestlings and fledglings; in others, such care is the responsibility of only one sex. In some species, other members of the same species-usually close relatives of the breeding pair, such as offspring from previous broods-will help with the raising of the young. Such alloparenting is particularly common among the Corvida, which includes such birds as the true crows, Australian Magpie and Fairy-wrens, but has been observed in species as different as the Rifleman and Red Kite. Among most groups of animals, male parental care is rare. In birds, however, it is quite common-more so than in any other vertebrate class. Though territory and nest site defence, incubation, and chick feeding are often shared tasks, there is sometimes a division of labour in which one mate undertakes all or most of a particular duty.
The point at which chicks fledge varies dramatically. The chicks of the Synthliboramphus murrelets, like the Ancient Murrelet, leave the nest the night after they hatch, following their parents out to sea, where they are raised away from terrestrial predators. Some other species, such as ducks, move their chicks away from the nest at an early age. In most species, chicks leave the nest just before, or soon after, they are able to fly. The amount of parental care after fledging varies; albatross chicks leave the nest on their own and receive no further help, while other species continue some supplementary feeding after fledging. Chicks may also follow their parents during their first migration.
Brood parasites
Brood parasitism, in which an egg-layer leaves her eggs with another individual's brood, is more common among birds than any other type of organism. After a parasitic bird lays her eggs in another bird's nest, they are often accepted and raised by the host at the expense of the host's own brood. Brood parasites may be either obligate brood parasites, which must lay their eggs in the nests of other species because they are incapable of raising their own young, or non-obligate brood parasites, which sometimes lay eggs in the nests of conspecifics to increase their reproductive output even though they could have raised their own young. One hundred bird species, including honeyguides, icterids, estrildid finches and ducks, are obligate parasites, though the most famous are the cuckoos. Some brood parasites are adapted to hatch before their host's young, which allows them to destroy the host's eggs by pushing them out of the nest or to kill the host's chicks; this ensures that all food brought to the nest will be fed to the parasitic chicks.
Ecology
Birds occupy a wide range of ecological positions. While some birds are generalists, others are highly specialised in their habitat or food requirements. Even within a single habitat, such as a forest, the niches occupied by different species of birds vary, with some species feeding in the forest canopy, others beneath the canopy, and still others on the forest floor. Forest birds may be insectivores, frugivores, and nectarivores. Aquatic birds generally feed by fishing, plant eating, and piracy or kleptoparasitism. Birds of prey specialise in hunting mammals or other birds, while vultures are specialised scavengers.
Some nectar-feeding birds are important pollinators, and many frugivores play a key role in seed dispersal. Plants and pollinating birds often coevolve, and in some cases a flower's primary pollinator is the only species capable of reaching its nectar.
Birds are often important to island ecology. Birds have frequently reached islands that mammals have not; on those islands, birds may fulfill ecological roles typically played by larger animals. For example, in New Zealand the moas were important browsers, as are the Kereru and Kokako today. Today the plants of New Zealand retain the defensive adaptations evolved to protect them from the extinct moa. Nesting seabirds may also affect the ecology of islands and surrounding seas, principally through the concentration of large quantities of guano, which may enrich the local soil and the surrounding seas.
Since birds are highly visible and common animals, humans have had a relationship with them since the dawn of man. Sometimes, these relationships are mutualistic, like the cooperative honey-gathering among honeyguides and African peoples such as the Borana. Other times, they may be commensal, as when species such as the House Sparrow have benefited from human activities. Several bird species have become commercially significant agricultural pests, and some pose an aviation hazard. Human activities can also be detrimental, and have threatened numerous bird species with extinction.
Birds can act as vectors for spreading diseases such as psittacosis, salmonellosis, campylobacteriosis, mycobacteriosis (avian tuberculosis), avian influenza (bird flu), giardiasis, and cryptosporidiosis over long distances. Some of these are zoonotic diseases that can also be transmitted to humans.
Economic importance
Domesticated birds raised for meat and eggs, called poultry, are the largest source of animal protein eaten by humans; in 2003, 76 million tons of poultry and 61 million tons of eggs were produced worldwide. Chickens account for much of human poultry consumption, though turkeys, ducks, and geese are also relatively common. Many species of birds are also hunted for meat. Bird hunting is primarily a recreational activity except in extremely undeveloped areas. The most important birds hunted in North and South America are waterfowl; other widely hunted birds include pheasants, wild turkeys, quail, doves, partridge, grouse, snipe, and woodcock. Muttonbirding is also popular in Australia and New Zealand. Though some hunting, such as that of muttonbirds, may be sustainable, hunting has led to the extinction or endangerment of dozens of species.
Other commercially valuable products from birds include feathers (especially the down of geese and ducks), which are used as insulation in clothing and bedding, and seabird feces (guano), which is a valuable source of phosphorus and nitrogen. The War of the Pacific, sometimes called the Guano War, was fought in part over the control of guano deposits.
Birds have been domesticated by humans both as pets and for practical purposes. Colourful birds, such as parrots and mynas, are bred in captivity or kept as pets, a practice that has led to the illegal trafficking of some endangered species. Falcons and cormorants have long been used for hunting and fishing, respectively. Messenger pigeons, used since at least 1 AD, remained important as recently as World War II. Today, such activities are more common either as hobbies, for entertainment and tourism, or for sports such as pigeon racing.
Amateur bird enthusiasts (called birdwatchers, twitchers or, more commonly, birders) number in the millions. Many homeowners erect bird feeders near their homes to attract various species. Bird feeding has grown into a multimillion dollar industry; for example, an estimated 75% of households in Britain provide food for birds at some point during the winter.
Religion, folklore and culture
Birds play prominent and diverse roles in folklore, religion, and popular culture. In religion, birds may serve as either messengers or priests and leaders for a deity, such as in the Cult of Makemake, in which the Tangata manu of Easter Island served as chiefs, or as attendants, as in the case of Hugin and Munin, two Common Ravens who whispered news into the ears of the Norse god Odin. They may also serve as religious symbols, as when Jonah (Hebrew: ??????, dove) embodied the fright, passivity, mourning, and beauty traditionally associated with doves. Birds have themselves been deified, as in the case of the Common Peacock, which is perceived as Mother Earth by the Dravidians of India. Some birds have also been perceived as monsters, including the mythological Roc and the Maori's legendary Pouakai, a giant bird capable of snatching humans.
Birds have been featured in culture and art since prehistoric times, when they were represented in early cave paintings. Birds were later used in religious or symbolic art and design, such as the magnificent Peacock Throne of the Mughal and Persian emperors. With the advent of scientific interest in birds, many paintings of birds were commissioned for books. Among the most famous of these bird artists was John James Audubon, whose paintings of North American birds were a great commercial success in Europe and who later lent his name to the National Audubon Society. Birds are also important figures in poetry; for example, Homer incorporated Nightingales into his Odyssey, and Catullus used a sparrow as an erotic symbol in his Catullus 2. The relationship between an albatross and a sailor is the central theme of Samuel Taylor Coleridge's The Rime of the Ancient Mariner, which led to the use of the term as a metaphor for a 'burden'. Other English metaphors derive from birds; vulture funds and vulture investors, for instance, take their name from the scavenging vulture.
Perceptions of various bird species often vary across cultures. Owls are associated with bad luck, witchcraft, and death in parts of Africa, but are regarded as wise across much of Europe. Hoopoes were considered sacred in Ancient Egypt and symbols of virtue in Persia, but were thought of as thieves across much of Europe and harbingers of war in Scandinavia.
Conservation
Though human activities have allowed the expansion of a few species, such as the Barn Swallow and European Starling, they have caused population decreases or extinction in many other species. Over a hundred bird species have gone extinct in historical times, although the most dramatic human-caused avian extinctions, eradicating an estimated 750-1800 species, occurred during the human colonisation of Melanesian, Polynesian, and Micronesian islands. Many bird populations are declining worldwide, with 1,221 species listed as threatened by Birdlife International and the IUCN in 2007. The most commonly cited human threat to birds is habitat loss. Other threats include overhunting, accidental mortality due to structural collisions or long-line fishing bycatch, pollution (including oil spills and pesticide use), competition and predation from nonnative invasive species, and climate change. Governments and conservation groups work to protect birds, either by passing laws that preserve and restore bird habitat or by establishing captive populations for reintroductions. Such projects have produced some successes; one study estimated that conservation efforts saved 16 species of bird that would otherwise have gone extinct between 1994 and 2004, including the California Condor and Norfolk Island Green Parrot.
Web Reference:
http://en.wikipedia.org/wiki/Bird
http://animals.nationalgeographic.com/animals/bugs.html
 Editor for Asisbiz: Matthew Laird Acred
Editor for Asisbiz: Matthew Laird Acred
If you love our website Please donate so we can make this site even better !!
Please donate so we can make this site even better !!